Camelid-derived single-domain antibodies, known as VHHs or nanobodies, are increasingly utilized in structural biology for their ability to bind diverse epitopes with high affinity and specificity. These recombinant fragments, approximately 15 kDa in size, derive from camelid antibodies and heavy-chain-only antibodies found in llamas, camels, and alpacas. Unlike classical antibodies, engineered fragments like scFvs, or conventional IgG antibodies, VHHs require no pairing between VH and VL domains and exhibit high solubility and stability due to their evolved framework mutations. The small VHH domain size and optimized amino acid sequence contribute to their stability and suitability for structural studies.
Their ability to access conformationally dynamic or hidden epitopes, high recombinant expression yields, and thermal stability make them particularly suitable for applications in structural biology, including X-ray crystallography and cryo-electron microscopy. Many researchers now use VHH antibody platforms to stabilize difficult membrane proteins and generate reproducible 3D structures for downstream analysis.
Biointron’s VHH Antibody Discovery Service offers custom immunization and phage display solutions to generate high-affinity VHHs optimized for structural studies. These workflows support the discovery of heavy chain variable domain antibodies with high antigen binding specificity across a wide range of targets.
The structural integrity and biochemical behavior of nanobodies are strongly influenced by their unique sequence and domain architecture. Key features include:
Extended CDR3 Loops: VHHs often contain longer complementarity-determining region 3 (CDR3) loops than conventional VH domains. These loops contribute to an expanded paratope surface area and can penetrate into concave or recessed protein surfaces inaccessible to conventional antibodies. The flexible side chains and unique amino acid repeat patterns within the CDR regions also contribute to recognition of hidden antigenic determinants and immunodominant epitopes.
Framework Region Adaptations: Mutations in framework region 2 (FR2) replace hydrophobic residues necessary for VL pairing in traditional antibodies with hydrophilic counterparts, enhancing solubility and preventing aggregation. These adaptations improve the biochemical stability of the llama VHH domain and reduce formation of protein aggregates during purification and crystallization workflows.
Increased Refolding Capacity: VHHs demonstrate high resilience to thermal denaturation and can refold without loss of antigen-binding capability, an asset in purification and crystallization workflows. These characteristics make VHH antibody constructs highly suitable for handling under challenging biochemical conditions.
These physicochemical properties make nanobodies suitable crystallization partners by reducing target flexibility and increasing conformational homogeneity. Structural biologists frequently combine surface plasmon resonance, thermal shift assays, and other biophysical methods to validate binding kinetics before structural studies.
Flexible proteins, such as membrane-bound receptors or disordered cytoplasmic domains, often resist crystallization due to structural heterogeneity. Nanobodies can stabilize these proteins by locking them into defined conformations. For example, Nb484 was used to stabilize the intrinsically disordered N-terminal domain of the human prion protein (PrP), allowing successful crystallization and revealing an extended β-sheet motif associated with amyloid plaques formation.
VHHs offer additional ordered surfaces that promote favorable crystal lattice formation. In studies of glycosylated human IL-23, co-crystallization with three distinct nanobodies masked flexible glycan regions, enabling the formation of diffraction-quality crystals and improved x-ray structure determination.
Nanobodies can distinguish between functionally distinct conformers, a property exploited to trap active or inactive states of receptors. Structural studies of the M2 muscarinic receptor utilized Nb9-8 to lock the receptor in an active-like state, promoting high-quality GPCR crystals. These approaches support advanced protein structure prediction and experimental validation workflows.
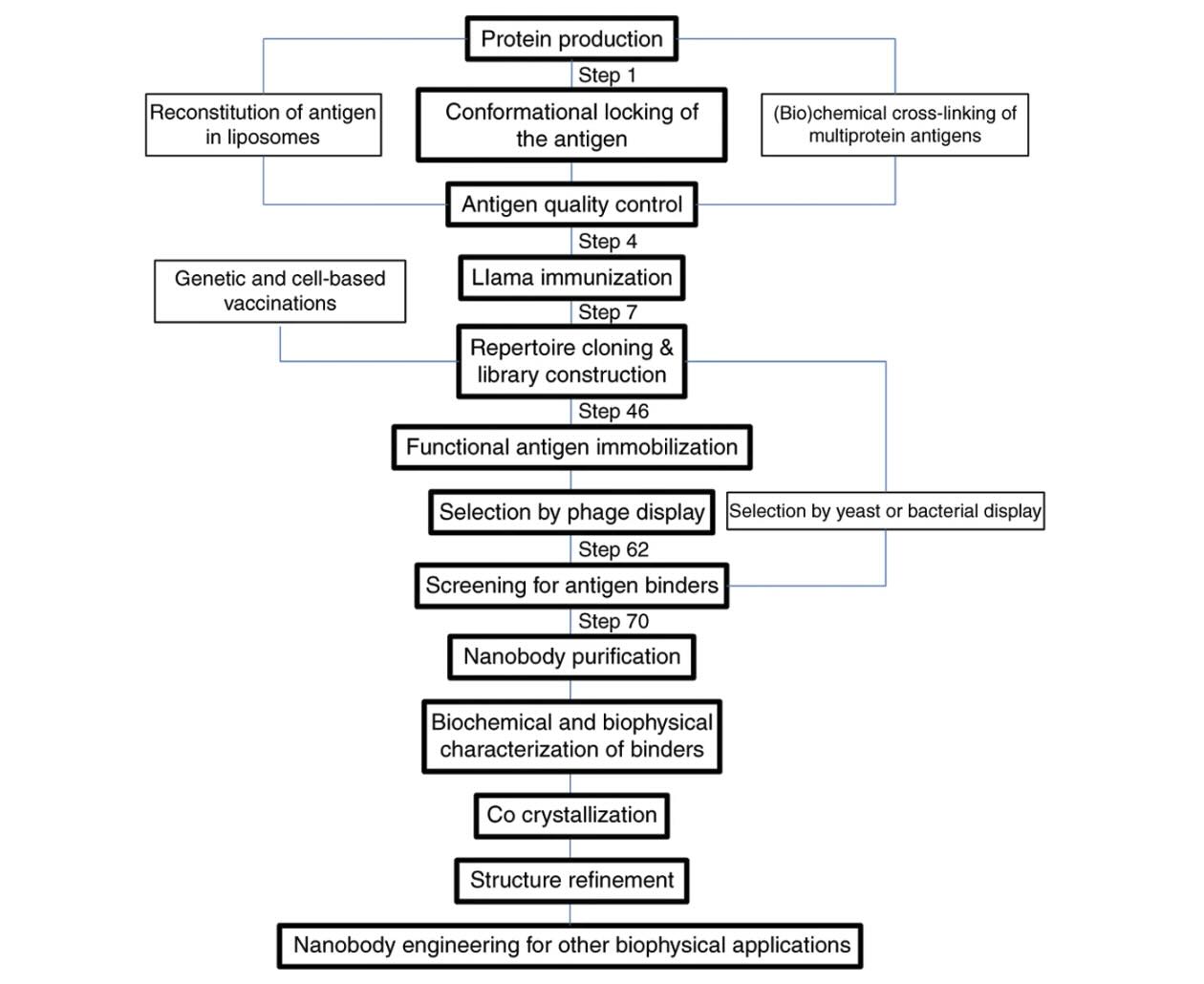
Nanobody selection for structural applications typically follows immunization of camelids from different camelid species with the target antigen. Lymphocyte-derived mRNA and other nucleic acid templates are reverse transcribed, and VHH fragments are amplified and cloned into phage display vectors and specialized expression vectors. Antigen-specific binders are selected through multiple rounds of panning using optimized biopanning protocol workflows and validated by ELISA, surface plasmon resonance, and complementary biophysical assays.
Naïve and synthetic VHH display libraries are also emerging as alternatives to immunization, although they may require more extensive screening. The growing availability of commercial libraries and transgenic platforms enables customized VHH discovery pipelines capable of generating both monovalent VHH and multivalent VHH constructs.
VHHs can be efficiently expressed in microbial systems, such as in E. coli and S. cerevisiae. These hosts support proper folding, disulfide bond formation, and scalable protein yields. Typical expression levels exceed 5–10 mg/L under optimized conditions. These systems are widely used for heterologous expression of recombinant antibody fragments and other engineered proteins.
Periplasmic targeting using signal peptides (e.g., PelB) enhances yield and simplifies purification. In cytoplasmic expression systems, VHHs remain soluble and functional due to their monomeric structure and lack of hydrophobic interfaces, unlike scFvs which often aggregate. The absence of a large fc domain further improves intracellular expression and solubility compared to conventional antibody formats.
Additionally, VHHs tolerate chemical denaturants and extreme pH, supporting downstream purification under a wide range of buffer conditions. Researchers commonly use western blotting, size exclusion chromatography (SEC), and analytical chromatography to validate protein integrity and confirm reproducibility of experimental data.
Despite their favorable properties, several factors must be considered when using VHHs in structural biology:
Target Interference: High-affinity binders may disrupt native protein-protein interactions or obscure functionally critical surfaces.
Orientation and Symmetry: Crystallization success depends on the geometry of the VHH-protein complex. Poor alignment may hinder lattice formation.
Epitope Selection: Conformationally selective binders must be chosen to avoid stabilizing non-physiological states or misfolded intermediates.
These challenges can be mitigated through rational binder selection and iterative co-crystallization trials, and computational structure prediction methods. Modern workflows increasingly integrate deep learning approaches such as AlphaFold2 predictions to improve binder design and optimize sequence identity comparisons across candidate VHHs.
Nanobodies are increasingly used in cryo-electron microscopy (cryo-EM) for complex stabilization and particle alignment. Their ability to lock flexible domains and bind buried epitopes enhances the quality of cryo-EM datasets, particularly in membrane protein studies. These advances are accelerating the generation of accurate VHH model systems for difficult targets.
Moreover, nanobodies can trace protein conformational dynamics in vitro and in live cells. They have been used to distinguish active and inactive states of EGFR, P-glycoprotein, and L-plastin, revealing mechanisms of signaling, transport, and immune synapse formation. Some imaging studies can combine green fluorescent protein, immunofluorescence microscopy, and nanobody labeling to visualize intracellular proteins such as lamins A and C in live-cell systems.
The ability to couple conformation-specific nanobodies with fluorescence tags further enables super-resolution imaging and live-cell structural biology. Modified constructs incorporating cell-penetrating peptides and secondary VHH Antibodies are also being explored for intracellular delivery and enhanced detection sensitivity.
For structural biology, the solubility, small size, and conformational selectivity of nanobodies have been exploited to:
Stabilize mobile domains in GPCRs, ion channels, and viral proteins
Improve crystallization of membrane-bound and glycosylated targets
Capture functionally relevant conformations for structure-guided drug design
Looking forward, the integration of VHHs with cryo-EM, NMR, and AI-driven modeling will further expand their role in protein structure determination. Computational pipelines using deep learning, protein structure prediction, and molecular simulations are expected to accelerate future VHH engineering workflows.
Biointron's VHH Antibody Discovery services are positioned to support these efforts through customized binder generation and expression workflows tailored to structural biology applications. These services support advanced studies involving immunoglobulin fragments, engineered bi-head VHH constructs, and emerging CAR binders for translational research applications.
What are nanobodies in structural biology?
Nanobodies are single-domain antibodies from camelids. They can be used to stabilize protein targets for crystallography and cryo-EM. These VHH domain binders are derived from naturally occurring heavy chain antibodies.
How do VHHs help in crystallography?
They can stabilize flexible proteins, mask mobile regions, and promote crystal lattice formation to improve X-ray structure resolution.
Are nanobodies used in cryo-EM?
Yes, they can improve particle homogeneity and enable trapping of distinct conformations in complex systems, such as in cryoelectron microscopy workflows.
How are VHHs produced for structural studies?
Recombinantly using systems such as E. coli, often using phage display technology after immunization or from synthetic display libraries.
Do VHHs interfere with protein function?
Some do. Structural validation ensures binding does not hinder native conformation or function while preserving relevant biological activity.
Yasaman Asaadi, Fatemeh Fazlollahi Jouneghani, Janani, S., & Fatemeh Rahbarizadeh. (2021). A comprehensive comparison between camelid nanobodies and single chain variable fragments. Biomarker Research, 9(1), 87–87. https://doi.org/10.1186/s40364-021-00332-6
Dmitriev, O. Y., Lutsenko, S., & Muyldermans, S. (2016). Nanobodies as Probes for Protein Dynamics in Vitro and in Cells. The Journal of biological chemistry, 291(8), 3767–3775. https://doi.org/10.1074/jbc.R115.679811
Desmyter, A., Spinelli, S., Roussel, A., & Cambillau, C. (2015). Camelid nanobodies: Killing two birds with one stone. Current Opinion in Structural Biology, 32, 1-8. https://doi.org/10.1016/j.sbi.2015.01.001
Explore VHH antibody production strategies for molecular imaging probes, includi……
Artificial intelligence and machine learning are expanding the number of antibod……
June 2026 included acquisitions of late-stage antibody developers, partnerships ……